Wanida Sukketsiri1 ![]() ,
Kitja Sawangjaroen1,
Supita Tanasawet2
,
Kitja Sawangjaroen1,
Supita Tanasawet2
For correspondence:- Wanida Sukketsiri Email: wanida.su@psu.ac.th Tel:+6674288171
Received: 24 December 2015 Accepted: 5 April 2016 Published: 27 May 2016
Citation: Sukketsiri W, Sawangjaroen K, Tanasawet S. Anti-apoptotic effects of phyllanthin against alcohol-induced liver cell death. Trop J Pharm Res 2016; 15(5):981-988 doi: 10.4314/tjpr.v15i5.12
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To evaluate the anti-apoptotic effect of phyllanthin on alcohol-induced liver cell death in HepG2 cells alone and in co-culture with human monocytic (THP-1) differentiated macrophage cells.
Methods: Cell viability was determined by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. The cells were pretreated with 1, 5 and 10 μM phyllanthin for 24 h followed by 1300 mM alcohol for HepG2 cells and 2000 mM alcohol for the co-cultured cells. Thereafter, intracellular reactive oxygen species (ROS), mitochondrial membrane potential (MMP) changes, apoptotic cell death and caspase-3/7 activities were assessed.
Results: Alcohol exposure significantly increased intracellular ROS generation (p < 0.001), decreased MMP changes (p < 0.001), increased the number of apoptotic and necrotic cells (p < 0.001) and also induced higher caspase-3/7 activity (p < 0.001) in the co-culture with THP-1 differentiated macrophage cells than in HepG2 cells alone. Pretreatment of HepG2 cells and co-cultured cells with phyllanthin for 24 h prior to alcohol exposure significantly decreased intracellular production of ROS (p < 0.001) and also increased the change in MMP (p < 0.001) as well as caused a decrease in the number of apoptotic and necrotic cells (p < 0.001), but inhibited caspase-3/7 activity (p < 0.001).
Conclusion: The results indicate that phyllanthin treatment may have a significant therapeutic effect on alcohol-related liver diseases.
Introduction
Alcohol consumption is a major cause of chronic liver disease which may develop after years of chronic alcohol consumption and results in high morbidity and mortality rates [1]. Moreover, hepatocyte apoptosis is significantly raised in alcohol hepatitis patients and correspond with the severity of the disease and hepatic fibrosis [2]. Alcohol-induced liver injury is a complex process and is dependent upon the interplay of multiple cell types in the liver. It is of interest, that hepatic macrophages (Kupffer cells) play a pivotal role in the pathogenesis of alcohol-induced liver cell death [3]. Kupffer cells are activated following alcohol consumption to generate various toxic mediators such as oxygen radicals, tumor necrosis factor alpha (TNF-α), interleukin and prostaglandins [4,5] contribute to hepatocyte dysfunction, apoptosis and the necrosis of hepatocytes [6,7].
Current alcohol liver disease therapy using modern medical practices is often restricted in its efficacy, has high adverse events and is costly [1]. So, there is a need to identify potential therapeutic agents that will lower the incidence of adverse effects. Therefore, medicinal plants are being used for alternative and complementary therapies. Preparations from Phyllanthus amaras Schum. et Thonn have been used extensively in traditional medicine particularly for the treatment of liver diseases [8]. In addition, phyllanthin, the active compound of P. amarus is reported to be an active antioxidant [9], anti-inflammatory [10,11] and hepatoprotective properties [9]. The mechanism of the hepatoprotective activity of phyllanthin has been shown to effectively facilitate the recovery of its antioxidant capability including the level of total glutathione, the activities of superoxide dismutase and glutathione reductase [12,13]. However, the effect of phyllanthin on alcohol-induced cell death has not been clarified.
Hence, the aim of this study was to investigate the efficacy of phyllanthin in preventing in vitro HepG2 alone and a co-culture of HepG2 with THP-1 differentiated macrophage cells after injury induced by exposure to alcohol.
Methods
Chemicals and cell culture
Phyllanthin was isolated and purified from the fresh aerial part of P. amarus and identified by IR spectrum and HPLC, as previously reported [12]. The purities of the phyllanthin preparations were compared with standard references (Chromadex, Inc., USA), and were at least 98 % pure [12]. THP-1 cells were obtained from the Cell Line Service, Heidelberg, Germany, and human hepatocarcinoma (HepG2) cells were kindly provided by Dr Wisit Tangkeangsirisin, Department of Biopharmacy, Faculty of Pharmacy, Silpakorn University, Thailand.
HepG2 cells were cultured in complete Dulbecco’s modified Eagle’s medium (DMEM) containing supplemented nutrients and essential condition (GIBCO, Invitrogen Corporation, USA). Before use, THP-1 cells were induced to differentiate into macrophage-like cells using 100 ng/mL of phorbol 12-myristate 13-acetate (Sigma-Aldrich, USA). The cells were cultured for 48 h to allow final differentiation into macrophages [14]. A co-culture of HepG2 and THP-1 differentiated macrophage cells at a ratio of 2:1 was grown in complete RPMI-1640 medium supplemented with nutrients and essential condition.
Cytotoxicity assay
HepG2 cells (1.5 x 104 cells/well) and co-cultured with THP-1 differentiated macrophage cells were treated with alcohol at concentrations of 0-2000 mM and further incubated for 4 h. In addition, both cell types were treated with phyllanthin at various concentrations (0-50 μM) for 24 h. Thereafter, cell viability was determined using a colorimetric MTT (Sigma-Aldrich, USA) assay [15]. Briefly, the MTT solution was added at a final concentration of 0.5 mg/mL and the optical density was measured at 570 nm using a microplate reader (Bio-tex, USA).
Intracellular reactive oxygen species (ROS) production
Intracellular ROS production in the HepG2 and co-culture cells were assayed through the oxidation of 2′,7′-dichlorofluorescein diacetate (DCFH-DA) (Sigma-Aldrich, USA) [16]. Briefly, both cell types were cultured at a density of 3 x 104 cells/well. Before alcohol exposure, cells were pretreated with phyllanthin at concentrations of 0, 1, 5 and 10 µM for 24 h. Then, cells were washed and incubated with 20 µM of DCFH-DA for 45 min followed by 1300 mM alcohol for the HepG2 cells and 2000 mM alcohol for the co-cultured cells for 4 h. The fluorescence intensity was measured using a fluorescence microplate reader (Bio-tex, USA) at an excitation wavelength of 485 nm and emission wavelength of 530 nm.
Mitochondrial membrane potential (MMP) analysis
The MMP change was analyzed using the MMP assay kit (Abcam, USA), according to the manufacturer’s instructions. In brief, both sets of cells (3 x 104 cells/well) were pretreated with phyllanthin (0-10 µM) for 24 h and then incubated with 1300 mM alcohol for the HepG2 cells and 2000 mM alcohol for the co-cultured cells for 4 h. The fluorescence intensity was measured using a fluorescence microplate reader (Bio-tex, USA) at an excitation wavelength of 549 nm and emission wavelength of 575 nm. Tetramethyl rhodamine ethyl ester (TMRE) positive cell was expressed as % of control fluorescence signal from the treated and untreated samples.
Apoptosis and necrosis assay
Apoptosis and necrosis were determined using a fluorescent DNA stain with Hoechst 33342 and propidium iodide (PI) (Cell Signaling, USA). The cells were stained with 10 μg/mL Hoechst 33342 and PI for 30 min at 37 oC. Cells were examined using a fluorescence microscope (Olympus IX71, Japan). The data was presented in terms of % cell death.
Evaluation of caspase-3/7 activity
Caspase-3/7 activity was measured using the Caspase-Glo assay kit (Promega, USA), according to the manufacturer’s instructions. Briefly, both of the cell types were cultured in a 96-well white plate at a density of 1 x 104 cells/well and pretreated with (0-10 µM) phyllanthin for 24 h, the cells were then treated with alcohol for 4 h. The luminescence intensity was measured using a luminometer (Applied Biosystems, USA) with parameters of a 1 minute lag time and 0.5 sec/well-read time.
Statistical analysis
All data are presented as a mean ± standard error of the mean (SEM, n = 4). One-way analysis of variance (ANOVA) with Tukey’s test was used to test differences in mean values among the groups. P < 0.05 was considered statistically significant. The data were analyzed using SPSS version 16.
Results
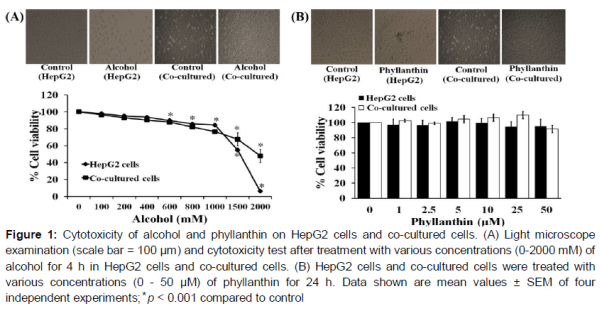
Cell morphology and cytotoxicity of phyllanthin and alcohol on HepG2 cells and co-cultured cells
Alcohol significantly decreased the viability of the HepG2 cells and the co-cultured cells in a concentration-dependent manner with morphological changes (the cells rounded up, lose contact with their neighbors and shrank) (A). The 50 % inhibitory concentrations (IC50) of alcohol in the HepG2 cells and co-cultured cells were 1225.42 ± 27.56 and 1950.20 ± 72.27 mM, respectively. In addition, our results indicated that phyllanthin (1-50 μM) by itself did not affect cell viability or morphology of any of the cell types after 24 h of treatment (B).
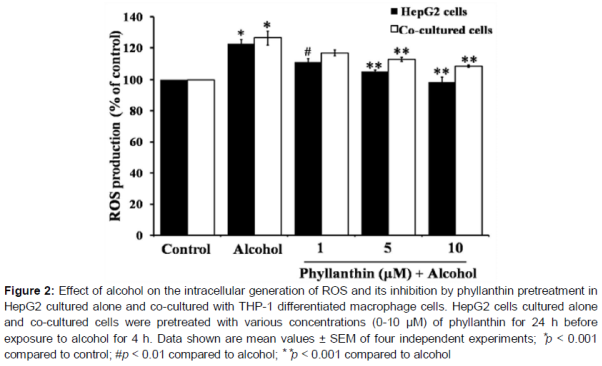
Effect of phyllanthin against alcohol-induced ROS production in HepG2 and co-cultured cells
Without phyllanthin pretreatment, alcohol produced a significant increase in the intracellular ROS production to 122.66 ± 2.56 and 126.43 ± 4.52 % of the control in the HepG2 and co-cultured cells, respectively after 4 h of alcohol exposure (). In HepG2 cells, we found that phyllanthin at a dose of 1, 5 and 10 µM could significantly attenuate ROS generation to 111.04 ± 2.27, 105.22 ± 0.92, and 98.50 ± 3.26 % of the control, respectively (). In the co-cultured cells, a high concentration of phyllanthin (5 and 10 µM) significantly decreased the intracellular ROS production to 112.89 ± 1.25 and 108.61 ± 0.77 % of the control, respectively (). In the hepatocytes cultured alone, these results indicated that alcohol caused an elevation in the intracellular ROS level; however, it was significantly reduced by phyllanthin pretreatment. On the other hand, only a high concentration of phyllanthin could prevent the alcohol-induced ROS generation in the co-cultured cells. Hence it might be the macrophages that induced the high ROS production.
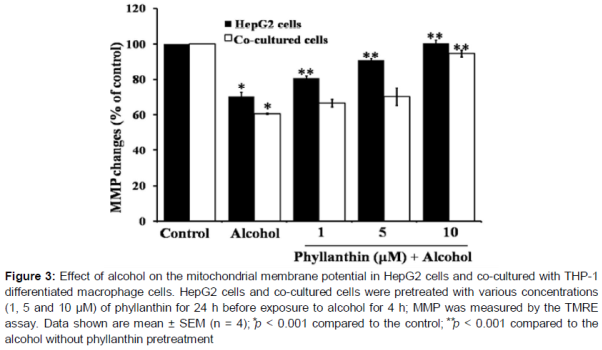
Effect of phyllanthin against alcohol-induced MMP change in HepG2 and co-cultured cells
Exposure to alcohol for 4 h significantly decreased the percentage of TMRE positive cells to 70.24 ± 2.31 and 60.52 ± 0.36 % of the control in HepG2 cells and the co-cultured cells, respectively (). The percentage of the TMRE positive cells was significantly increased in a concentration-dependent manner when the cells were pretreated with (1 - 10 μM) phyllanthin before alcohol exposure of the HepG2 cells (). In the co-cultured cells, pretreatment of phyllanthin only at its highest concentration (10 μM) significantly increased the percentage of TMRE positive cells (). It can be inferred from the obtained results that phyllanthin could significantly reduce the alcohol-induced mitochondrial dysfunction.
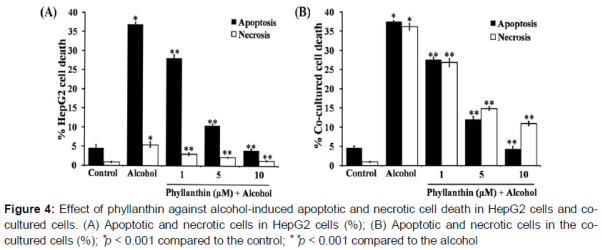
Effect of phyllanthin against alcohol-induced apoptosis in HepG2 and co-cultured cells
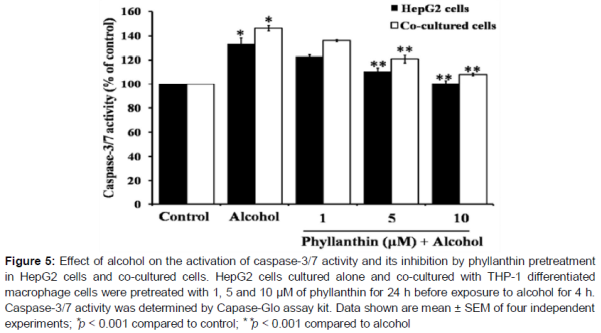
Alcohol exposure significantly increased in the percentage of apoptotic and necrotic cells in the HepG2 cells (A). In addition, there was a significant increase in the caspase-3/7 activation when compared with the untreated cells (). Surprisingly, exposure to a high concentration of alcohol showed a stronger activity to diminish the viable cells by inducing apoptosis and necrotic cell death in the co-cultured cells than HepG2 (B and ). These data indicated that macrophages were involved in the severity of alcohol-induced liver cell death. However, phyllanthin at 1, 5 and 10 μM caused a decrease in the percentage of apoptotic and necrotic cells (A and 4B) and also significantly inhibited caspase-3/7 activation by alcohol in both HepG2 cells and the co-cultured cells (). These results indicated that phyllanthin had an anti-apoptotic activity against alcohol-induced toxicity in both cell types. Taken together, these finding indicated that phyllanthin attenuated necrosis in both cell types.
Discussion
Phyllanthin has anti-oxidative activity, anti-inflammatory activity [9,10,17] and protects against alcohol-induced liver injury via its antioxidant properties [12]. Our MTT assay showed that alcohol exposure significantly reduced cell viability and altered the morphology (the cells rounded up, lose contact with their neighbors and shrank) of the HepG2 single cell cultures and also when co-cultured with THP-1 differentiated macrophage cells and that this corresponded to the increase in intracellular ROS production. Alcohol-induced oxidative stress plays a major role in the mechanism by which alcohol is involved in liver injury [18] and it triggers cellular damage in various tissues including the brain, liver and retina [12,19,20].
The advancement of alcohol-induced liver injury involves both parenchymal and non-parenchymal cells of the liver. Hence macrophages could make hepatocytes more susceptible to alcohol injury by increasing the production of ROS [21]. However, pretreatment with phyllanthin prevented the production of ROS induced by alcohol in both HepG2 cells and co-cultured cells. This result is consistent with a previous finding of decreased oxidative stress to chemical-induced toxicity of hepatocyte cells after phyllanthin treatment [9,12]. In addition, phyllanthin had a strong inhibitory activity on the increase in production of nitric oxide radical by macrophages [10] and oxidative burst in neutrophils [22]. The residues produced by ROS caused a lowering of the MMP leading to an uncontrolled ROS formation and release of proapoptotic factors such as cytochrome c into the cytosol where they promote activation of caspase [23,24] and subsequently apoptosis.
In the present study, alcohol caused a significant decrease in the MMP that led to a more profound reduction in the co-cultured cells than the HepG2 culture alone, presumably because of the involvement of ROS in the destruction of the MMP. Also, this finding is consistent with a previous observation that acute alcohol treatment induced apoptosis in primary hepatocytes accompanied by ROS production and mitochondrial depolarization [25,26]. In the current study, pretreatment with phyllanthin impeded the decrease of MMP in the HepG2 cells and the co-cultured cells. This can partly explain the hepatoprotective potential of phyllanthin that may maintain the normal redox status and improve mitochondrial function.
Cells undergoing apoptosis present several characteristic morphological and molecular changes [27]. Chromatin condensations, membrane blebbing, phosphatidylserine externalization, DNA fragmentation and, eventually, the disruption of the cell into membrane-bound fragments are some of the distinct morphological and biochemical features [28] associated with apoptosis.
Acute alcohol exposure promotes apoptosis in primary hepatocyte cultures that was linked with ROS formation, mitochondrial depolarization, and cytochrome c release [25]. In this study, alcohol exposure induced a higher severity of liver toxicity by both apoptotic and necrotic cell death in co-culture with THP-1 differentiated macrophage cells than in HepG2 cells alone. Macrophage cells may also, in part, be involved in the severity of alcohol-induced co-cultured cell death. Several reports have indicated that there is a relationship between Kupffer cells and the severity of alcohol-induced liver damage [29,30].
After being activated, macrophage cells secrete a category of cytokines such as TNF-α, interleukin one beta and thus the activated macrophage cells are the main source of ROS in the liver [5,29-31]. These inflammatory mediators lead to hepatocyte dysfunction, apoptosis and necrosis of hepatocytes, and the production of extracellular matrix proteins contributing to characteristic fibrosis [30,31].
Considering these results, pretreating cells with phyllanthin was effective in inhibiting alcohol-induced cell death by inhibiting caspase-3/7 activation and also by decreasing the number of apoptotic and necrotic cells in the HepG2 cells alone and the co-cultured cells. Meanwhile, a P. amarus extract decreased DNA fragmentation in acetic acid-induced colitis in rats [11]. In addition, there is some evidence that phyllanthin inhibited TNF-α expression and prevented activation of the nuclear factor-κB in hepatic tissue [32]. Thus, the finding in this study clearly support the anti-apoptotic role of phyllanthin, the mechanism of which was to ameliorate the disturbances caused by oxidative stress and loss of mitochondrial function, with inhibition of the subsequent apoptosis cascade.
Conclusion
Alcohol-induced liver cell death is caused by increased ROS generation, decreased MMP and induced caspase-3/7 activation in both HepG2 cells and those co-cultured with THP-1 differentiated macrophage cells. Phyllanthin prevents alcohol-induced cell death in human liver cells by decreasing the production of ROS and increasing MMP change and as well as by inhibiting the activation of caspase-3/7 activity. These results suggest that phyllanthin can be developed for hepatoprotective therapy against alcohol-induced liver disease.
Declarations
Acknowledgement
References
Archives


News Updates